Liigi teke ja spetsiatsioon: kuidas uued liigid tekivad
Kui uus liik tekib, millised mehhanismid seda käivitavad — anagenees, kladogenees, geeniülekanne ja hübriidistumine. Sügav ülevaade liigi tekkest ja spetsiatsioonist.
Spetsiatsioon ehk liigi teke käsitleb seda, kuidas liigid tekivad ja muutuvad uute, geneetiliselt eristuvate rühmadeni. See on evolutsioonibioloogia keskne teema, mis selgitab, miks ja kuidas elusolendid jagunevad uute taksonoomiliste üksusteni, omandades püsivad erinevused väljanägemises, käitumises ja genoomis. Spetsiatsiooni uurimisel kasutatakse nii morfoloogilisi, käitumuslikke kui ka molekulaarseid andmeid (DNA järjestuse analüüs) ning seda seostatakse tihedalt evolutsioonibioloogiaga.
Pildigalerii
10 Pildid









Ajalooline taust ja põhimõisted
Darwin pakkus, et uued liigid tekivad järkjärgulise muutumise kaudu olemasolevatest liikidest – seda protsessi nimetatakse anageneesiks (liigi sees toimuv pikaajaline muutumine ehk "fütogenees"). 20. sajandi suurem osa rõhku asetati aga kladogeneesile: liigi jagunemisele kaheks või enamaks eri haruks, kus ühest eelsugulasest tekivad paralleelselt mitu uut liiki. Kladogeneesi kirjeldamisel on oluline roll isoleeruvatel mehhanismidel, mis piiravad geenivoogu ja soodustavad geneetilist lahknemist.
Peamised spetsiatsiooni tüübid
- Allopaatiline spetsiatsioon – liigid tekivad siis, kui populatsioonid jäävad geograafiliselt eraldatuks (näiteks mäestiku, jõe või mandri serva tõttu). Eraldatus takistab regulaarset paaritumist ja võimaldab kromosoomide, geenide ja omaduste sukeldumist eri suundades.
- Peripaatiline spetsiatsioon – sarnaneb allopaatilisele, kuid uus liik tekib tavaliselt väikese, asurkonna äärealal oleva asutajarühma põhjal; siin võivad mängida suurt rolli asutajaefekt ja geneetiline drift.
- Parapaatiline spetsiatsioon – toimub siis, kui populatsioonid asuvad üksteisele ligidal, kuid elavad eri elupaikade graadil; osaline geenivoog toimub, kuid keskkonnatingimuste erinevused suunavad kohastumist ja divergeerumist.
- Sümpaatiline spetsiatsioon – uued liigid tekivad samal geograafilisel alal ilma nähtava füüsilise barjäärita. Sellisel juhul võivad spetsiatsiooni käivitada tugev eristuv valik, erinev toitumisharjumus, eri ajastusel paaritumine või kromosoomsete muutuste (nt. polüploidia) tõttu tekkiv kiire isolatsioon.
Isolatsiooni mehhanismid ja nende roll
Spetsiatsiooni eelduseks on enamasti mingisugune isolatsioon, mis vähendab või katkestab geenivoogu järgmiste peamiste mehhanismide kaudu:
- Eelhübriidne isolatsioon (prezygotic) – takistused, mis ennetavad paaritumist või viljastamist: ruumiline/ajaliselt eristumine, käitumuslikud erinevused (erinev laul või lõhnaaineid kasutav paaritumiskäitumine), reproduktiivorganite sobimatus või gametide mittesobivus.
- Järelhübriidne isolatsioon (postzygotic) – kui hübriidid tekivad, on neil sageli vähenenud ellujäämine või viljakus (nt. hambutu või steriliseeritud järglased), mis vähendab geenide liikumist liigi vahel.
Geneetilised ja ökoloogilised põhjused
Spetsiatsioon ei ole pelgalt geograafiline protsess; olulised on ka geneetilised jõud ja ökoloogiline eristumine. Peamised tegurid on:
- Looduslik valik – erinevad keskkonnatingimused valivad erinevaid omadusi, mis võivad aja jooksul viia püsiva eristumiseni.
- Sexuaalne valik – paaritumueelistused võivad kiirendada geneetilist lahknemist (nt eri värvuse või laulu eelistus).
- Geneetiline drift ja asutajaefekt – eriti väikestes populatsioonides võivad juhuslikud muutused geenifrekventis põhjustada kiiret erinevust.
- Kromosoomsete muutuste mõju – polüploidia on eriti tähtis taimede puhul: kahe eri liigi ristumine ja järgnev kromosoomide dubleerumine võib kiiresti luua uue, reproduktiivselt eraldunud rühma.
Hübriidimine ja introgressioon
Viimase paari aastakümne uurimistöö ning DNA järjestuse võrdlus on näidanud, et sugulasliikide vahel esineb sageli hübriidistumist. Hübriidid võivad kanda edasi geene ühest liigist teise – seda nimetatakse introgressiooniks. See tuletab meelde, et reproduktiivne isolatsioon ei pea olema täielik, et eristumine toimuks; liigid võivad jagada geene ja siiski säilitada eristatavaid tunnuseid. Mõnikord viib hübriidimine hoopis uue liigi tekkeni (hübriidspetsiatsioon), tõsiste näidetega taimedel (nt mitmed õistaimede liigid) ja loomadel (näiteks mõnedes päevaliblikate ja päevalugude rühmades).
Praktilised näited
- Darwini tihased Galapagose saartel – allopaatiline spetsiatsioon ja kohastumine erinevate toitumisallikate suhtes.
- Ensatina-värviplaastrad (mõnes sisemaise salamandri rühmas) – näide ringliigi fenomeenist, kus naaberpopulatsioonid võivad paljuneda, kuid “otsa” populatsioonid on eraldunud.
- Aafrika järvede cichlidae – kiire ökoloogiline spetsiatsioon, kus toit ja käitumine on toonud kaasa suurt liigirikkust.
- Taimede polüploidia (näiteks genus Tragopogon) – kiire, tihti hübriidne liigi teke kromosoomide dubleerimise kaudu.
Ajaskaala ja spetsiatsiooni olemus
Spetsiatsioon võib võtta ajaliselt väga erineva kestuse: mõnel juhul (nt polüploidia) võib uus liik tekkida ühe või paari põlvkonnaga; teinekord võib see protsess võtta kümneid tuhandeid kuni miljoneid aastaid. Oluline on mõista, et spetsiatsioon on protsess – üleminek ühest pidevast geneetilisest variatsioonist mitmeks püsivaks, eristuvaks rühmaks. On olemas spetsiatsiooni „lõikeline“ ja „järkjärguline“ mudel ning reaalsus sisaldab sageli mõlema elemente.
Kokkuvõte
Spetsiatsioon on kompleksselt juhitud protsess, kus geograafiline eraldatus, geneetilised muutused, looduslik ja seksuaalne valik, ökoloogiline eristumine ning vahel ka hübriidimine ja kromosoomsed mutatsioonid mängivad olulist rolli. Kuigi kunagine rõhk oli peamiselt füüsilisel eraldatusel (allopaatia), on tänased vaatlused ja DNA-analüüsid näidanud, et liigi teke on mitmekesine ja et reproduktiivne isolatsioon ei pea olema absoluutselt täielik, enne kui uued liigid tekivad.
Isolatsioonimehhanismid
Isolatsioonimehhanismid on asjad, mis takistavad liigi rühmade vahelist edukat paljunemist. Populatsioonide paljunemisalane isolatsioon on kehtestatud. See on eriti oluline bioloogilise liigi mõiste jaoks, kuna liigid on määratletud reproduktiivse isolatsiooni kaudu.
Isolatsioonimehhanismid võib jagada kahte rühma, enne ja pärast viljastamist.
Enne viljastamist
Tegurid, mis takistavad isendite paaritumist.
- Geograafiline isolatsioon: Liigid esinevad eri piirkondades ja on sageli eraldatud tõkete abil.
- Ajaline isolatsioon: Isendid ei paaritu, sest nad on aktiivsed erinevatel aegadel. See võib olla eri kellaaegadel või erinevatel aastaaegadel. Liikide paaritumisajad ei pruugi kokku langeda. Isendid ei kohtavad üksteist kas nende paaritumisperioodidel või üldse mitte.
- Ökoloogiline isolatsioon: Isendid paarituvad ainult oma eelistatud elupaigas. Nad ei puutu kokku teiste liikide isenditega, kellel on teistsugused ökoloogilised eelistused.
- Käitumuslik isolatsioon: Erinevatest liikidest isendid võivad kohtuda, kuid üks neist ei tunnista mingeid seksuaalseid vihjeid, mida võidakse anda. Indiviid valib enamasti oma liigi liikme.
- Mehaaniline isolatsioon: Kopulatsiooni võib püüda, kuid spermasiiret ei toimu. Isendid võivad olla suuruse või morfoloogia tõttu kokkusobimatud.
- Gameetiline kokkusobimatus: Spermasiirde toimub, kuid muna ei viljastu.
Pärast viljastamist
Tegurid, mis takistavad paaritumise õnnestumist, näiteks geneetiline kokkusobimatus, hübriidide eluvõimetus või steriilsus.
- Zügootiline suremus: Muna viljastub, kuid tsütot ei arene.
- Hübriidi eluvõimetus: Hübriidembrüo moodustub, kuid ei ole elujõuline.
- Hübriidsteriilsus: Hübriid on elujõuline, kuid saadud täiskasvanud isend on steriilne.
- Hübriidne jaotus: Esimese põlvkonna (F1) hübriidid on elujõulised ja viljakad, kuid edasised hübriidipõlvkonnad (F2 ja tagasiristamised) on elujõuetu või steriilne.

Geograafiline isolatsioon
See on arvatavasti kõige tavalisem põhjus, mis põhjustab liigi tekkimist. Esimene inimene, kes selle peale mõtles, oli Moritz Wagner, saksa maadeuurija ja loodusteadlane.
Wagner alustas oma karjääri geograafina ja avaldas mitmeid geograafilisi raamatuid Põhja-Aafrika, Lähis-Ida ja Troopilise Ameerika kohta. Ta oli ka innukas loodusteadlane ja kollektsionäär ning just selle töö poolest on ta bioloogide seas kõige tuntum. Evolutsionist ja bioloogiaajaloolane Ernst Mayr on andnud ülevaate Wagneri tähtsusest. lk 562-565
Kolme Alžeerias veedetud aasta jooksul uuris Wagner (muude tegevuste hulgas) lennuvõimetuid mardikaid Pimelia ja Melasoma. Kumbki perekond jaguneb mitmeks liigiks, millest igaüks on piiratud Atlase mägedest Vahemerre laskuvate jõgede vahelise põhjaranniku osaga. Niipea, kui üks neist ületab jõe, ilmub teine, kuid lähedalt suguluses olev liik.
"... algav liik [tekib] ainult siis, kui mõned üksikud indiviidid ületavad oma levikuala piirid... uue rassi moodustamine ei õnnestu kunagi... ilma kolonistide pikaajaliseta eraldatuseta oma liigi teistest liikmetest".
See oli varajane kirjeldus ühe geograafilise liigi muutumise protsessist. Mayr võttis selle 1942. aastal uuesti kasutusele ja geograafilise speciatsiooni tähtsusest sai üks evolutsioonilise sünteesi keskseid ideid.
Teine termin geograafilise speciatsiooni kohta on allopatriline speciatsioon. Allopatria tähendab "eri maad".
Vabad saared
Vulkaanilised saared tekivad ilma eluta ja kogu elu peab saabuma tuule või vee abil. Hawaii saarte ja Galapagose saarte kohta teame, et kõik eluvormid muutuvad, kui nad jõuavad mandrilt saartele.
Hawaii saartel on umbes 6 500 ruutmiilil (17 000 km2) kõige mitmekesisem drosofiilide kärbeste kogum maailmas, mis elab vihmametsadest kuni mäginiitudeni. On teada umbes 800 Hawaii drosofiilide liiki.
Uuringud näitavad selget liikide "voolu" vanematelt saartelt uuematele saartele. On ka juhtumeid koloniseerimisest tagasi vanematele saartele ja saarte vahelejätmisest, kuid need on palju harvemad.
Kaaliumi/argooni radioaktiivse dateerimise järgi pärinevad praegused saared 0,4 miljonit aastat tagasi (mya) (Mauna Kea) kuni 10mya (Necker). Hawaii saarestiku vanim veel mere kohal olev liige on Kure atoll, mida saab dateerida 30 mya.
Saarestik ise, mis on tekkinud Vaikse ookeani plaadi liikumisel üle kuuma koha, on eksisteerinud palju kauem, vähemalt kriidiajastul. Hawaii saared pluss endised saared, mis on nüüd mere all, moodustavad Hawaii-Emperori merealuse mäeahela; ja paljud veealused mäed on tüübimäed.
Kõik Hawaii kohalikud drosofiilide liigid pärinevad ilmselt ühest esivanemaliigist, mis koloniseeris saared umbes 20 miljonit aastat tagasi. Järgnevat kohanemisradiatsiooni kiirendas konkurentsi puudumine ja mitmesugused vabad niššid. Kuigi oleks võimalik, et saare koloniseeris üks tiinus emasloom, on tõenäolisem, et tegemist oli samast liigist pärit rühmaga.
Hawaii saarestikus on ka teisi loomi ja taimi, mis on läbinud sarnase, kui ka vähem suurejoonelise kohanemisradiaatori.
Rõngaliigid
Bioloogias on rõngaliigid seotud rida naaberpopulatsioone, millest igaüks võib ristuda naaberpopulatsioonidega. Kaks ahela otsa kattuvad.
Sarja kaks lõpupopulatsiooni on liiga kaugel suguluses, et nad saaksid omavahel ristuda. Sellised mittesugunevad, kuigi geneetiliselt seotud "lõpp-populatsioonid" võivad samas piirkonnas koos eksisteerida, sulgedes seega "rõnga".
Rõngaliigid on olulised tõendid evolutsiooni kohta: nad illustreerivad, mis juhtub aja jooksul, kui populatsioonid geneetiliselt lahknevad. Richard Dawkins täheldas, et rõngaliigid "näitavad meile ainult ruumilises dimensioonis midagi, mis peab alati toimuma ajalises dimensioonis".
Siiski on raske leida lihtsat ja otsest näidet.
Larus kajakad
Klassikaline näide ringliikide kohta on Larus lokkide tsirkumpolaarne liigirõngas. Nende kajakate levikuala moodustab ümber põhjapooluse rõnga, mida üksikud kajakad tavaliselt ei lenda üle.
Laias laastus võivad kajakad oma naabritega mingil määral ristuda, välja arvatud rõnga kahes otsas.
Väike- ja heeringalokid on piisavalt erinevad, et nad tavaliselt ei hübriideeru; seega moodustab lokkide rühm järjepidevuse, välja arvatud seal, kus need kaks sugukonda Euroopas kohtuvad.
Kuigi tegelikkus on palju keerulisem, näitab see hästi üldist ideed.
Ensatina salamandrid
Ensatina salamander on rõngaliigiks Kalifornia keskse oru ümbruse mägedes. Kompleks moodustab ümber mägede hobuseraua kuju. Kuigi iga 19 hobuseraua ümber asuva populatsiooni vahel võib toimuda ristumine, ei saa hobuseraua läänepoolses otsas asuv Ensatina eschscholtzii alamliik ristuda idapoolses otsas asuva Ensatina klauberi alamliigiga. See on näide "peaaegu kõikidest liigirikkuse etappidest" (Dobzhansky). Richard Highton väitis, et Ensatina puhul on tegemist mitme liigi, mitte ühe liigi järjepidevusega.
Roheliseks muutuv laulurähn (The Greenish Warbler)
Rohekaarel (Phylloscopus trochiloides) on mitu alamliiki, millest P. t . viridianus on Euroopas kõige tuntum. Tegemist on rõngaliigiga, mille populatsioonid lahknevad Tiibeti platoo ida- ja läänepoolses osas, kohtudes hiljem põhjapoolses osas. Nende sugulussuhted on üsna segased.




Sümpatriline liigiline mitmekesisus
Sümpatriline liikumine tähendab kahe või enama järglasliigi moodustumist ühest esivanemaliigist, mis kõik asuvad samas geograafilises piirkonnas. Seda peetakse praegu väga sagedaseks.
Sümpatrilise liigirikkuse puhul erinevad liigid, kui nad elavad samas kohas. Sageli viidatud näiteid sümpatrilisest liigirikkusest on leitud putukate puhul, kes muutuvad sõltuvaks erinevatest peremeestaimedest samas piirkonnas.
Sümpatrilise speciatsiooni kui liigirikkuse mehhanismi olemasolu oli teravalt vaidlustatud. Väideti, et sümpatrilise liigirikkuse tõendid on tegelikult mikrogeograafilise liigirikkuse näited. Üldiselt ei ole see praegu eelistatud seletus. Pigem on see lähedaste sugulasliikide vahelise hübriidistumise tulemus, millele järgneb looduslik valik, mis mõjub selliste ristandite järglastele. Üks laialdaselt tunnustatud näide sümpatrilisest liigilisusest on Ida-Aafrikas asuva Nabugabo järve tsichliidide puhul, mille põhjuseks peetakse sugulist valikut.
Spetsialiseerumine polüploidsuse kaudu
Polüploidsus on põhjustanud palju kiireid liigirikkuse sündmusi, sest näiteks tetraploidse x diploidse paarumise järglased annavad sageli triploidse steriilse järeltulija.
Siiski ei ole kõik polüploidid oma vanemtaimedest reproduktiivselt isoleeritud ja geenivool võib siiski toimuda näiteks triploidsete hübriidide ja diploidide paaritumise kaudu, mis annavad tetraploide.
Paljud olemasolevad taime- ja enamik loomaliike on ilmselt oma evolutsioonilise ajaloo jooksul läbinud polüploidiseerumise. Edukate polüploidsete liikide paljunemine on mõnikord aseksuaalne, parthenogeneesi teel. Teadmata põhjustel on paljud aseksuaalsed organismid polüploidsed.
Hawthorn fly
Üks näide evolutsiooni toimimisest on raagoletis pomonella, mis näib olevat läbimas sümpatrilist liigilist muutumist.
Erinevad orasekärbse populatsioonid toituvad erinevatest viljadest. Eraldi populatsioon tekkis Põhja-Ameerikas 19. sajandil, mõni aeg pärast seda, kui sisse toodi õunad, mis ei ole kohalik liik. See õunapopulatsioon toitub tavaliselt ainult õunadest, mitte aga ajalooliselt eelistatud oraseviljadest. Praegune orasrohu toitev populatsioon ei söö tavaliselt õunu.
Mõned tõendid viitavad sellele, et toimub sümpatriline liigiline areng. Puuvõõrikukärbsed küpsevad hiljem ja nende suguküpsus võtab kauem aega kui õunakärbse; samuti on vähe tõendeid ristumise kohta (teadlased on dokumenteerinud 4-6% hübriidsuse määra).
Uue orasekärbse teke on näide käimasolevast evolutsioonist.
Hübriidimine
Harva tekib uus liik, kui eri liikide üksikud liikmed paarituvad. Tavaliselt on selliste hübriidpaaride saadused viljatu (mitte viljakad) või suhteliselt viljatu ja seega kõrvaldatakse need loodusliku valiku teel.
On leitud üks näide uue ja eduka hübriidliigi kohta. Uus liik on Itaalia ja Hispaania varblaste hübriid, mille põhitõdesid on kontrollitud nende vere DNA järjestusanalüüsi abil. Lind elab Itaalias piirkonnas, kus elavad mõlemad vanemliigid. Ta ei paljune koos hispaania varblasega, kuigi elab nendega kõrvuti.
Tehislik liigitamine
Uusi liike on loodud koduloomade kasvatamise teel, kuid esialgsed kuupäevad ja meetodid on ebaselged. Näiteks kodulambad on loodud hübriidistamise teel ja nad ei anna enam elujõulisi järeltulijaid Ovis orientalis'e, ühest liigist, millest nad põlvnevad.
Koduveiseid võib seevastu pidada samaks liigiks mitmete metshärgade, gauride, jakide jne. liikidega, kuna nad annavad nendega kergesti viljakat järeltulijat.
Laboratooriumi liigid
Kõige paremini dokumenteeritud uute liikide loomine laboris toimus 1980. aastate lõpus. William Rice ja G. W. Salt aretasid puuviljakärbseid, Drosophila melanogaster, kasutades labürinti, kus oli kolm erinevat elupaigavalikut, nagu valgus/pime ja märg/kuiv.
Iga põlvkond paigutati labürinti ja kaheksast väljapääsust kahest väljapääsust väljunud kärbeste rühmad paigutati eraldi, et nad omavahel oma rühmades paljuneksid. Pärast kolmkümmend viis põlvkonda olid kaks rühma ja nende järeltulijad reproduktiivselt isoleeritud, sest nad eelistasid elupaiku tugevalt: nad paaritusid ainult nende poolt eelistatud aladel ja seega ei paaritunud kärbeste vahel, kes eelistasid teisi alasid. Selliste katsete ajalugu on kirjeldatud Rice ja Hostert (1993).
Diane Dodd suutis samuti näidata, kuidas Drosophila pseudoobscura paaritumis-eelistustest võib areneda reproduktiivne isolatsioon juba kaheksa põlvkonna järel, kasutades erinevaid toidutüüpe, tärklist ja maltoosi.
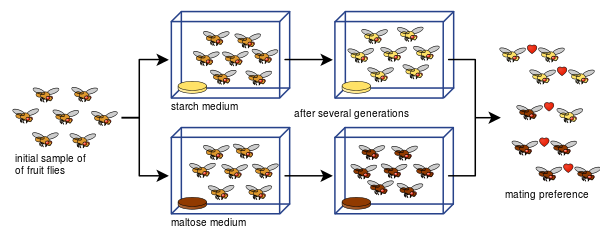
Doddi eksperimenti on olnud lihtne korrata paljude teiste poolt, sealhulgas teiste puuviljakärbeste ja toiduga.
Tugevdamine (Wallace'i efekt)
Tugevdamine on protsess, mille abil looduslik valik suurendab reproduktiivset isolatsiooni.
See võib tekkida pärast seda, kui kaks sama liigi populatsiooni eraldatakse ja seejärel taas kokku puutuvad. Kui nende reproduktiivne isolatsioon oli täielik, siis on neist juba kujunenud kaks erinevat kokkusobimatut liiki.
Kui nende reproduktiivne isolatsioon ei ole täielik, siis tekivad edasised paaritumised populatsioonide vahel hübriidid, mis võivad olla viljakad või mitte. Kui hübriidid on viljatu või viljakad, kuid vähem sobivad kui nende esivanemad, siis ei toimu edasist reproduktiivset isolatsiooni ja liigiline eristumine on sisuliselt toimunud (nt nagu hobuste ja eeslite puhul).
Seda põhjendatakse sellega, et kui hübriidsete järglaste vanematel on kummalgi looduslikult valitud tunnused oma eri keskkondade jaoks, siis kannab hübriidsed järglased mõlema omadusi ja ei sobi kummalegi niššile sama hästi kui kumbki vanem. Hübriidide vähene sobivus põhjustaks valiku, mis soodustaks assortatiivset paaritumist, mis vähendaks hübriidistumist.
Seda nimetatakse mõnikord Wallace'i efektiks, lähtudes evolutsioonibioloog Alfred Russel Wallace'ist, kes 19. sajandi lõpus pakkus välja, et see võib olla oluline tegur liigirikkuse puhul.
Kui hübriidide järeltulijad on sobivamad kui nende esivanemad, siis sulanduvad populatsioonid kokkupuutepiirkonnas tagasi samaks liigiks.
Tugevdamine on vajalik eraldamiseks, kui kahe liigi vormi vahel on "hübriidvöönd". Hübriiditsoonid on piirkonnad, kus erinevad populatsioonid kohtuvad ja ristuvad. Hübriidsed järeltulijad on nendes piirkondades väga levinud, mis tavaliselt tekivad lahknevate liikide teisejärgulise kokkupuute tulemusel.
Ilma tugevdamiseta oleks kahe populatsiooni või liigi ristumine kontrollimatu. Tugevdamist võib esile kutsuda kunstliku valiku katsetes, nagu eespool kirjeldatud.
Küsimused ja vastused
K: Mis on erisus?
V: Spetsifikatsioon on protsess, mille käigus tekivad liigid. See on oluline osa evolutsioonibioloogiast.
K: Mida arvas Darwin liikide tekkimisest?
V: Darwin arvas, et enamik liike tekkis otse juba olemasolevatest liikidest, mida ta nimetas anageneesiks või "fütoloogiliseks evolutsiooniks".
K: Kuidas vaadeldi 20. sajandil üldiselt liikide teket?
V: 20. sajandil arvasid teadlased, et enamik liike tekkis varasemate liikide jagunemisel, mida nimetati kladogeneesiks. Samuti arvati, et seda jagunemist põhjustasid või aitasid kaasa isoleerivad mehhanismid, näiteks füüsiline eraldumine.
K: Kuidas on hiljutised teadusuuringud muutnud meie arusaama liigilisusest?
V: Hiljutised uuringud on näidanud, et sugulasliikide vahel võib toimuda hübriidistumine ja nende ristamiste kaudu võivad geenid edasi kanduda. See tähendab, et reproduktiivne isolatsioon ei ole alati vajalik liigi määratlemiseks ja et liigi tekkimiseks ei pruugi olla vaja allopatriat (füüsilist eraldumist).
K: Kuidas on füüsiline eraldatus seotud uute liikide moodustumisega?
V: Arvatakse, et füüsiline eraldumine kunagi koos eksisteerinud liikide vahel on peamine tegur uute liikide tekkimisel, mida illustreerivad paljud teaduskirjanduses käsitletud näited.
Seotud artiklid
Autor
AlegsaOnline.com Liigi teke ja spetsiatsioon: kuidas uued liigid tekivad Leandro Alegsa
URL: https://et.alegsaonline.com/art/92570
Allikad
- doi.org : 10.1126/science.23.587.506
- pubmed.ncbi.nlm.nih.gov : 17789700
- doi.org : 10.1086/279001
- science.siu.edu : "The concept of species"
- abacus.gene.ucl.ac.uk : "Isolating mechanisms"
- mallet.oeb.harvard.edu : mallet.oeb.harvard.edu/publications/what-speciation
- books.google.com : "The Continental Drift Controversy: Introduction of Seafloor Spreading," p. 292
- pubs.usgs.gov : Volcanism in Hawaii, U.S. Geological Survey Professional Paper 1350, pp. 5-54
- rspb.royalsocietypublishing.org : "The herring gull complex is not a ring species"
- doi.org : 10.1098/rspb.2004.2679
- journals.royalsoc.ac.uk : Electronic Appendix
- pnas.org : Incipient species formation in salamanders of the Ensatina complex
- jstor.org : "Is Ensatina eschscholtzii a ring-species?"
- doi.org : 10.1146/annurev.genet.34.1.401
- pubmed.ncbi.nlm.nih.gov : 11092833